Arkea
Arkea atau Archaea adalah domain mikroorganisme bersel satu. Arkea tergolong prokariota, kelompok mikrob yang tidak memiliki inti sel dan organel yang dibatasi membran. Awalnya, arkea diklasifikasikan sebagai bakteri dan diberi nama archaebacteria (di kerajaan Archaebacteria), tetapi klasifikasi ini tak lagi digunakan. Sel arkea memiliki sifat unik yang memisahkan arkea dari dua domain kehidupan yang lain, yaitu Bacteria dan Eukaryota. Arkea dibagi lagi menjadi beberapa filum. Klasifikasinya sulit dilakukan karena mayoritas arkea belum diisolasi di laboratorium dan hanya dideteksi urutan gennya dari sampel yang diambil dari lingkungan. Arkea dan bakteri umumnya memiliki ukuran dan bentuk yang serupa, meskipun beberapa arkea memiliki bentuk yang sangat aneh, misalnya sel-sel datar dan berbentuk persegi seperti Haloquadratum walsbyi. Meskipun terdapat kesamaan morfologi antara arkea dengan bakteri, arkea memiliki gen-gen dan beberapa jalur metabolik yang lebih serupa dengan eukariota, terutama enzim yang terlibat dalam transkripsi dan translasi. Salah satu aspek biokimia arkea yang unik adalah ketergantungan arkea pada lipid eter di membran sel mereka. Arkea menggunakan lebih banyak sumber energi dibandingkan eukariota, yang berkisar dari senyawa organik seperti gula, amonia, ion logam, atau bahkan gas hidrogen. Arkea yang toleran terhadap garam (Haloarchaea) menggunakan sinar matahari sebagai sumber energi, sementara spesies arkea lain memfiksasi karbon; namun, tidak seperti tumbuhan dan sianobakteri, tidak ada arkea yang melakukan kedua cara tersebut. Arkea bereproduksi secara aseksual dengan pembelahan biner, fragmentasi, atau tunas. Tidak seperti bakteri dan eukariota, tidak ada spesies arkea yang diketahui membentuk endospora. Arkea pertama yang diamati merupakan ekstremofil yang hidup di lingkungan ekstrem, seperti mata air panas dan danau garam tanpa organisme lain. Meningkatnya kemampuan alat deteksi molekuler memungkinkan penemuan arkea di hampir semua habitat, termasuk tanah, lautan, dan rawa. Jumlah mereka sangat banyak di lautan, dan arkea dalam bentuk plankton mungkin merupakan salah satu kelompok organisme yang paling melimpah di planet ini. Arkea berperan besar dalam kehidupan di Bumi. Mereka merupakan bagian dari mikrobiota bagi semua organisme. Sebagai mikrobiota manusia, arkea berperan penting di usus besar, rongga mulut, dan kulit. Keragaman morfologis, metabolik, dan geografis memungkinkan mereka memiliki peran ekologis yang luas, misalnya pada fiksasi karbon dalam siklus karbon, peran dalam siklus nitrogen, perputaran senyawa organik, dan memelihara komunitas simbiosis dan sintrofi mikrob. Belum ditemukan contoh yang jelas untuk arkea yang bersifat patogenik atau parasitik. Sebaliknya, arkea sering kali bersifat mutualis atau komensal, misalnya metanogen yang hidup di saluran pencernaan manusia dan ruminansia, tempat mereka hidup dalam jumlah besar dan membantu pencernaan. Metanogen digunakan dalam produksi biogas dan pengolahan limbah. Sementara itu, enzim dari arkea ekstremofil yang dapat bertahan pada suhu tinggi dan pelarut organik dimanfaatkan dalam bioteknologi. KlasifikasiKonsep awal.jpg) Pada hampir sepanjang abad ke-20, prokariota dianggap sebagai satu kelompok organisme dan diklasifikasikan berdasarkan sifat biokimia, morfologi, dan metabolisme mereka. Para ahli mikrobiologi mencoba mengklasifikasikan mikroorganisme berdasarkan struktur dinding sel, bentuk, dan zat-zat yang mereka konsumsi.[6] Pada tahun 1965, Linus Pauling dan Emile Zuckerkandl[7] mengusulkan untuk menggunakan urutan gen dalam berbagai prokariota untuk mengetahui hubungan mereka satu sama lain. Pendekatan ini, yang dikenal sebagai filogenetik, adalah metode utama yang digunakan saat ini.[8] Arkea (saat itu hanya metanogen yang diketahui) pertama kali diklasifikasikan sebagai kelompok yang terpisah dari bakteri pada pohon filogenetik berdasarkan urutan gen RNA ribosomal (rRNA) mereka oleh Carl Woese dan George E. Fox pada tahun 1977.[9] Awalnya, kedua kelompok tersebut masing-masing diberi nama Archaebacteria dan Eubacteria, dan diperlakukan sebagai kerajaan atau subkerajaan, yang oleh Woese dan Fox disebut Urkingdoms. Namun, klasifikasi ini tak lagi digunakan.[10] Woese dan Fox berpendapat bahwa Archaebacteria secara fundamental berbeda dari kehidupan yang lain: (1) ketiadaan peptidoglikan pada dinding sel mereka; (2) adanya dua koenzim yang tidak biasa; dan (3) hasil pengurutan gen RNA ribosomal 16S. Untuk menekankan perbedaan-perbedaan ini, Woese, Otto Kandler, dan Mark Wheelis kemudian mengusulkan sistem pengelompokan organisme yang baru dengan tiga domain terpisah: Eukarya, Bacteria, dan Archaea,[1] yang sekarang dikenal sebagai "Revolusi Woesian".[11] Kata arkea berasal dari bahasa Yunani Kuno ἀρχαῖα, yang berarti "hal-hal kuno",[12] karena wakil pertama dari domain Archaea adalah metanogen dan diasumsikan bahwa metabolisme mereka mencerminkan suasana primitif Bumi dan mereka merupakan organisme kuno. Untuk waktu yang lama, arkea terlihat sebagai ekstremofil yang hanya ada di habitat ekstrem seperti mata air panas dan danau garam. Namun, seiring bertambahnya habitat baru yang dipelajari, semakin banyak arkea yang ditemukan. Mikrob-mikrob halofilik ekstrem[13] dan hipertermofilik[14] juga termasuk dalam Archaea. Pada akhir abad ke-20, arkea juga telah diidentifikasi di lingkungan nonekstrem. Saat ini, mereka dikenal sebagai kelompok organisme yang besar dan beragam yang tersebar luas di alam.[15] Arkea mendapatkan apresiasi baru mengenai peranan dan keberadaan mereka sejak reaksi berantai polimerase (PCR) digunakan untuk mendeteksi prokariota dari sampel lingkungan (seperti air atau tanah) dengan melipatgandakan gen ribosom mereka. Hal ini memungkinkan deteksi dan identifikasi organisme yang belum dibudidayakan di laboratorium.[16][17] Klasifikasi Klasifikasi arkea, dan prokariota pada umumnya, adalah bidang yang bergerak cepat dan diperdebatkan. Sistem klasifikasi saat ini bertujuan untuk mengatur arkea ke dalam kelompok organisme yang memiliki karakteristik struktural dan nenek moyang yang sama.[18] Klasifikasi ini sangat bergantung pada pengurutan gen RNA ribosomal untuk mengungkapkan hubungan di antara berbagai organisme, yang dikenal sebagai filogenetik molekuler.[19] Sebagian besar spesies arkea yang dapat dikultur dan diteliti dengan baik adalah anggota dari dua filum utama, Euryarchaeota dan Crenarchaeota. Kelompok-kelompok lain dibuat secara tentatif. Misalnya, spesies aneh Nanoarchaeum equitans, yang ditemukan pada tahun 2003 dan telah diberikan filum sendiri, Nanoarchaeota.[20] Korarchaeota juga telah diusulkan sebagai filum baru yang berisi sekelompok kecil spesies termofilik yang berbagi karakteristik dengan kedua filum utama, tetapi lebih berkerabat dekat dengan Crenarchaeota.[21][22] Spesies arkea lain ternyata berkerabat jauh dengan salah satu dari kelompok ini, seperti Archaeal Richmond Mine acidophilic nanoorganisms (ARMAN) yang ditemukan pada tahun 2006[23] dan merupakan salah satu organisme terkecil yang diketahui.[24] Sebuah superfilum, yang disebut TACK, yang mencakup Thaumarchaeota, Aigarchaeota, Crenarchaeota, dan Korarchaeota, diusulkan pada 2011 dan mungkin terkait dengan organisme yang menjadi moyang eukariota.[25] Pada tahun 2017, superfilum Asgard telah diajukan sebagai kelompok yang lebih dekat dengan eukariota dan merupakan kelompok saudari dari TACK.[26] KladogramBerikut ini adalah kladogram arkea berdasarkan Tom A. Williams dkk. (2017)[27] serta Castelle dan Banfield (2018)[28] (DPANN):
Konsep spesiesKlasifikasi spesies-spesies arkea juga kontroversial. Biologi mendefinisikan spesies sebagai sekelompok organisme yang saling terkait. Kriteria perkembangbiakan eksklusif yang diketahui secara umum (organisme dalam satu spesies dapat berkembang biak dengan satu sama lain tetapi tidak dengan yang lain) tidak membantu karena arkea bereproduksi secara aseksual.[32] Arkea menunjukkan tingkat transfer gen horizontal yang tinggi di antara garis keturunan mereka. Beberapa peneliti menyarankan agar sejumlah individu dikelompokkan ke dalam populasi-populasi yang menyerupai spesies, yang genom-genomnya sangat mirip dan jarang mentransfer gen atau menerima transfer gen dari sel lain yang genomnya kurang terkait, seperti dalam genus Ferroplasma.[33] Di sisi lain, studi terhadap Halorubrum menemukan adanya transfer genetik yang signifikan ke/dari populasi yang kurang terkait, sehingga penerapan kriteria ini terbatas.[34] Kekhawatiran kedua adalah sejauh mana sebutan spesies memiliki arti praktis.[35] Pengetahuan terkini tentang keanekaragaman genetik masih tidak lengkap dan jumlah spesies arkea tidak dapat diperkirakan dengan akurat.[19] Perkiraan jumlah filum berkisar antara 18–23, tetapi hanya delapan filum yang memiliki perwakilan yang telah dikultur di laboratorium dan dipelajari secara langsung. Banyak kelompok-kelompok arkea dihipotesiskan hanya dari urutan rRNA tunggal, yang menunjukkan bahwa perbedaan di antara organisme ini tetap tidak jelas.[36] Bakteri juga mengandung banyak mikrob yang belum dikultur sehingga karakterisasi mereka juga serupa dengan arkea.[37] Asal-usul dan evolusiUmur Bumi diperkirakan sekitar 4,54 miliar tahun.[38][39][40] Bukti ilmiah menunjukkan bahwa kehidupan di Bumi dimulai setidaknya 3,5 miliar tahun yang lalu.[41][42] Bukti-bukti awal kehidupan di Bumi adalah grafit yang bersifat biogenik pada batuan metasedimentari berumur 3,7 miliar tahun yang ditemukan di Greenland Barat[43] dan fosil tikar mikrob yang ditemukan di batu pasir setua 3,48 miliar tahun di Australia Barat.[44][45] Pada tahun 2015, kemungkinan "sisa-sisa kehidupan biotik" ditemukan di batuan berumur 4,1 miliar tahun di Australia Barat.[46][47] Meskipun fosil sel prokariota kemungkinan berasal dari hampir 3,5 miliar tahun yang lalu, sebagian besar prokariota tidak memiliki morfologi yang dapat dibedakan, dan bentuk fosil tidak dapat digunakan untuk mengidentifikasi mereka sebagai arkea.[48] Sebaliknya, fosil kimia lipid yang unik menjadi lebih informatif karena senyawa tersebut tidak ada pada organisme lain.[49] Beberapa publikasi menyatakan bahwa sisa-sisa lipid arkea atau eukariota yang hadir dalam batu serpih berasal dari 2,7 miliar tahun yang lalu;[50] meskipun data tersebut dipertanyakan.[51] Lipid tersebut juga terdeteksi di batuan yang bahkan lebih tua dari batuan di Greenland Barat. Jejak tertua berasal dari distrik Isua, Greenland, yang termasuk sedimen tertua yang dikenal di Bumi yang terbentuk 3,8 miliar tahun yang lalu.[52] Silsilah arkea mungkin menjadi garis keturunan yang paling kuno yang ada di Bumi.[53] Woese berpendapat bahwa bakteri, arkea, dan eukariota mewakili garis keturunan yang terpisah yang bercabang sejak awal dari koloni leluhur semua organisme.[54][55] Salah satu kemungkinannya[55][56] adalah bahwa percabangan ini terjadi sebelum evolusi sel, saat tidak adanya membran sel memungkinkan transfer gen lateral yang tak terbatas, dan bahwa nenek moyang dari tiga domain tersebut muncul dari fiksasi subset-subset gen yang spesifik.[55][56] Ada kemungkinan bahwa nenek moyang terakhir dari bakteri dan arkea adalah termofil, sehingga suhu yang lebih rendah mungkin saja merupakan "lingkungan yang ekstrem" bagi arkea, dan organisme-organisme yang hidup di lingkungan dingin baru muncul belakangan.[57] Karena kekerabatan Archaea dan Bacteria lebih berjauhan satu sama lain dibandingkan antara mereka dengan eukariota, istilah prokariota mungkin menunjukkan kesamaan yang salah antara arkea dan bakteri.[58] Perbandingan dengan domain lainTabel berikut membandingkan beberapa karakteristik utama dari tiga domain, untuk menggambarkan persamaan dan perbedaan mereka.[59]
Archaea dipisahkan sebagai domain ketiga karena memiliki perbedaan besar dalam struktur RNA ribosomal mereka. Molekul RNA tertentu yang dikenal sebagai RNA ribosomal 16S (biasanya disingkat menjadi 16s rRNA) adalah kunci penting dalam produksi protein di semua organisme. Karena fungsi ini sangat penting dalam kehidupan, organisme yang mengalami mutasi 16s rRNA tidak mungkin bertahan hidup sehingga struktur nukleotida ini menjadi sangat stabil (tetapi tidak absolut) selama beberapa generasi. Ukuran 16s rRNA juga cukup besar untuk menyimpan informasi spesifik pada organisme tertentu, tetapi cukup kecil untuk diurutkan dan dibandingkan dalam waktu singkat. Pada tahun 1977, Carl Woese, seorang ahli mikrobiologi yang mempelajari urutan genetik organisme, mengembangkan metode perbandingan baru yang melibatkan pemecahan RNA menjadi fragmen-fragmen yang bisa diurutkan dan dibandingkan dengan fragmen lain dari organisme lain.[9] Semakin mirip pola fragmen tersebut di antara spesies-spesies, hubungan kekerabatannya menjadi lebih dekat.[62] Woese menggunakan metode perbandingan rRNA barunya untuk mengelompokkan dan memisahkan organisme yang berbeda. Ia mengurutkan berbagai spesies dan menemukan bahwa sekelompok metanogen dengan rRNA ternyata sangat berbeda dibandingkan prokariota atau eukariota yang telah dikenal.[9] Metanogen ini jauh lebih mirip satu sama lain dibandingkan dengan organisme lain yang pernah diurutkan, sehingga Woese mengusulkan domain baru, yaitu Archaea.[9] Eksperimen Woese menunjukkan bahwa arkea lebih mirip dengan eukariota alih-alih prokariota, meskipun struktur mereka lebih mirip dengan prokariota.[63] Hal ini menyimpulkan bahwa Archaea dan Eukarya memiliki nenek moyang bersama yang lebih baru dibandingkan dengan Eukarya dan Bacteria.[63] Perkembangan inti sel terjadi setelah percabangan antara bakteri dan nenek moyang bersama ini.[63] Salah satu sifat arkea yang unik adalah penggunaan lipid dengan ikatan eter secara berlimpah di membran sel mereka. Secara kimiawi, ikatan eter lebih stabil dari ikatan ester yang ditemukan dalam Bacteria dan Eukarya, yang mungkin menjadi faktor yang berkontribusi terhadap kemampuan banyak arkea untuk bertahan di lingkungan ekstrem, yang memberikan stres berat pada membran sel, seperti panas ekstrem dan salinitas. Analisis komparatif genom arkea juga telah mengidentifikasi beberapa indel khusus yang terkonservasi secara molekuler dan protein tertentu yang secara unik ditemukan baik di semua arkea maupun ditemukan di beberapa kelompok-kelompok utama dalam arkea .[64][65][66] Sifat lain yang unik dari arkea adalah bahwa tidak ada organisme lain yang dikenal mampu melakukan metanogenesis (produksi metana melalui metabolisme). Arkea metanogen memainkan peran penting dalam ekosistem bersama dengan organisme yang memperoleh energi dari oksidasi metana (kebanyakan dari mereka adalah bakteri) karena mereka sering menjadi sumber utama metana di lingkungan tersebut dan dapat berperan sebagai produsen primer. Metanogen juga memainkan peran penting dalam siklus karbon, memecah karbon organik menjadi metana, yang juga merupakan gas rumah kaca utama.[67] Hubungan dengan bakteri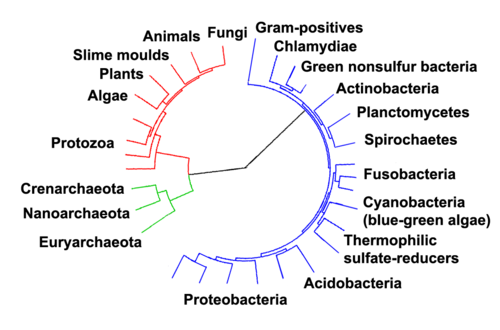 Hubungan di antara tiga domain sangat penting untuk memahami asal-usul kehidupan. Sebagian besar lintasan metabolisme (yang merupakan objek dari sebagian besar gen suatu organisme) sama-sama dimiliki Archaea dan Bacteria, sementara sebagian besar gen yang terlibat dalam ekspresi genom sama-sama dimiliki Archaea dan Eukarya.[69] Di antara prokariota, struktur sel arkea paling mirip dengan bakteri Gram-positif, terutama karena keduanya memiliki lipida dwilapis tunggal[70] dan biasanya mengandung sakulus tebal (eksoskeleton) dari berbagai komposisi kimia.[71] Pada beberapa pohon filogenetik yang didasarkan pada urutan gen-gen (protein-protein) dari homolog prokariota, homolog arkea lebih berhubungan dekat dengan homolog bakteri Gram-positif.[70] Arkea dan bakteri Gram-positif juga berbagi indel yang dilestarikan dalam sejumlah protein penting, seperti Hsp70 dan glutamin sintetase I;[70][72] namun, ilmuwan menafsirkan filogeni dari gen-gen ini sebagai sarana untuk mentransfer gen interdomain,[73][74] dan mungkin tidak mencerminkan hubungan kekerabatan organisme.[75] Ada pendapat bahwa arkea berevolusi dari bakteri Gram-positif sebagai respons terhadap tekanan seleksi antibiotik.[70][72][76] Pandangan ini diajukan berdasarkan pengamatan bahwa arkea tahan terhadap berbagai antibiotik yang terutama dibuat untuk bakteri Gram-positif,[70][72] dan bahwa antibiotik-antibiotik ini terutama bekerja pada gen yang membedakan arkea dari bakteri. Pendapat ini menyatakan bahwa tekanan selektif terhadap resistansi yang dihasilkan oleh antibiotik Gram-positif akhirnya cukup untuk menyebabkan perubahan yang luas pada banyak gen target antibiotik, dan bahwa galur ini mewakili nenek moyang arkea masa kini.[76] Evolusi arkea dalam menanggapi pemilihan antibiotik, atau tekanan selektif kompetitif lain, juga bisa menjelaskan adaptasi mereka terhadap lingkungan yang ekstrem (seperti suhu tinggi atau keasaman) sebagai hasil dari pencarian relung kosong untuk melarikan diri dari organisme yang memproduksi antibiotik;[76][77] Cavalier-Smith juga membuat usulan yang sama.[78] Pendapat ini juga didukung oleh studi lainnya yang menyelidiki hubungan protein struktural[79] dan studi yang menunjukkan bahwa bakteri Gram-positif mungkin merupakan awal garis keturunan yang bercabang paling awal dalam prokariota.[80] Hubungan dengan eukariotaHubungan evolusi antara arkea dan eukariota masih belum jelas. Selain dari kesamaan dalam struktur dan fungsi sel yang dibahas di bawah, banyak pohon genetik menyatukan keduanya dalam satu kelompok.[81] Faktor-faktor yang memperrumit hubungan mereka termasuk klaim bahwa hubungan antara eukariota dan filum arkea Crenarchaeota lebih dekat daripada hubungan antara filum Crenarchaeota dan Euryarchaeota,[82] dan adanya gen-seperti-arkea pada bakteri tertentu, seperti pada Thermotoga maritima, dari transfer gen horizontal.[83] Hipotesis standar menyatakan bahwa nenek moyang eukariota berpisah dari Archaea sejak awal,[84][85] dan bahwa eukariota muncul melalui fusi antara arkea dan bakteri, yang menjadi inti sel dan sitoplasma; hal ini menjelaskan berbagai kesamaan genetik tetapi kesulitan dalam menjelaskan struktur sel.[82] Sebuah hipotesis alternatif, hipotesis eosit, berpendapat bahwa Eukaryota muncul relatif terlambat dari Archaea.[86] Sebuah garis keturunan arkea yang ditemukan pada 2015, Lokiarchaeum (dari filum baru Lokiarchaeota), yang dinamai berdasarkan sebuah ventilasi hidrotermal yang disebut Kastel Loki di Samudra Arktik, ternyata merupakan kelompok yang paling berkerabat dekat dengan eukariota. Organisme ini disebut sebagai organisme peralihan antara prokariota dan eukariota.[87][88] Beberapa filum saudara dari "Lokiarchaeota" telah ditemukan ("Thorarchaeota", "Odinarchaeota", "Heimdallarchaeota"), yang semuanya membentuk sebuah supergrup Asgard yang baru diajukan, yang mungkin merupakan takson saudara dari Proteoarchaeota.[5][26][89] Rincian dari hubungan anggota Asgard dan eukariota masih dalam pertimbangan,[90] meskipun, pada Januari 2020, para ilmuwan melaporkan bahwa Candidatus Prometheoarchaeum syntrophicum, sejenis arkea Asgard, mungkin merupakan penghubung antara mikroorganisme prokariota yang sederhana dan mikroorganisme eukariota yang kompleks sekitar dua miliar tahun yang lalu.[91][92] MorfologiSatu individu arkea memiliki diameter yang berkisar dari 0,1 mikrometer (μm) sampai lebih dari 15 μm, yang hadir dalam berbagai bentuk, umumnya sebagai bola, batang, spiral, atau piring.[93] Morfologi lain ditemukan pada filum Crenarchaeota, termasuk sel berlobus yang bentuknya tidak teratur pada genus Sulfolobus, filamen seperti-jarum yang diameternya kurang dari setengah mikrometer pada genus Thermofilum, serta batang yang bentuknya hampir persegi panjang sempurna pada Thermoproteus dan Pyrobaculum.[94] Haloquadratum walsbyi adalah arkea dengan sel berbentuk persegi yang datar, yang hidup di kolam hipersalin.[95][96] Bentuk yang tidak biasa ini mungkin dipertahankan dengan baik oleh dinding sel dan sitoskeleton prokariotik mereka. Protein yang terkait dengan komponen sitoskeleton organisme lain ditemukan di arkea,[97] dan filamen-filamen terbentuk dalam sel mereka,[98] tetapi berbeda dengan organisme lain, struktur seluler ini kurang dipahami.[99] Dalam Thermoplasma dan Ferroplasma, ketiadaan dinding sel berarti bahwa sel memiliki bentuk yang tidak teratur, dan dapat menyerupai ameba.[100] Beberapa spesies arkea membentuk agregat atau filamen sel yang panjangnya hingga 200 μm.[93] Organisme-organisme ini bisa membentuk tonjolan dalam biofilm.[101] Khususnya, agregat sel-sel Thermococcus coalescens menyatu bersama dalam kultur sehingga membentuk sel-sel raksasa tunggal.[102] Genus Pyrodictium menghasilkan koloni multisel rumit yang melibatkan tabung berongga panjang dan tipis yang disebut kanula yang menonjol dari permukaan sel dan menghubungkan mereka ke dalam aglomerasi padat seperti-semak.[103] Fungsi dari kanula ini tidak disepakati, tetapi mereka dapat memungkinkan komunikasi atau pertukaran nutrisi dengan tetangganya.[104] Koloni multispesies juga ditemukan, seperti komunitas "untaian mutiara" yang ditemukan pada tahun 2001 di sebuah rawa di Jerman. Koloni bulat keputihan dari spesies baru Euryarchaeota terletak di sepanjang filamen tipis yang panjangnya dapat mencapai 15 sentimeter (5,9 inci); filamen ini terbuat dari spesies bakteri tertentu.[105] Struktur, perkembangan komposisi, dan operasiArkea dan bakteri memiliki struktur sel yang umumnya sama, tetapi komposisi dan organisasi sel membedakan arkea dengan bakteri. Seperti bakteri, arkea tidak memiliki membran interior dan organel.[58] Membran sel arkea, juga seperti bakteri, biasanya dibatasi oleh dinding sel dan mereka "berenang" menggunakan satu flagela atau lebih.[106] Secara struktural, arkea paling mirip dengan bakteri Gram-positif. Kebanyakan arkea memiliki membran plasma dan dinding sel tunggal, tanpa ruang periplasmik; pengecualian untuk aturan umum ini adalah Ignicoccus, yang memiliki sebuah periplasma sangat besar yang berisi vesikel bermembran dan tertutup oleh membran luar.[107] Dinding sel dan flagelaKebanyakan arkea (kecuali Thermoplasma dan Ferroplasma) memiliki dinding sel.[100] Mayoritas dinding sel arkea dirakit dari protein lapisan permukaan, yang membentuk lapisan-S.[108] Lapisan-S adalah susunan molekul protein yang kaku yang menutupi bagian luar sel (seperti zirah rantai).[109] Lapisan ini memberi perlindungan kimia dan fisik, serta dapat mencegah | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
